





Citation: Hudec I, Mock A (2014) Niphargus plurispinosus sp. n. (Crustacea, Amphipoda), a stygophile and hypotelminorheic representative from Central Europe. Subterranean Biology 13: 65–87. doi: 10.3897/subtbiol.13.6531
The detailed description of the morphology of Niphargus plurispinosus sp. n. from Slovakia is presented. Over 300 specimens were collected from a permanent seepage spring on repeated visits between May 2011 and May 2013. The type locality is located in the foothills of the Zemplínske vrchy mountains in the East Slovakian Lowland (NE part of Pannonian Lowland) - small, low and isolated hills formed during Neogene volcanic activity. Volcanic rocks draw together fragments of massives of Palaezoic and Mesozoic age as same as Neogene sediments. The new species can be classified as stygophile, living in the shallow subterranean habitat. The species has small subequal gnathopods, sexually dimorphic uropod III, sexually non-dimorphic uropod I in juveniles, dimorphic uropod 1 in adults. They are extremely different in the post-reproductive stage, when they have 2-4 dorsal spines (arranged in a transverse row) on the telson and supporting dorsal spines.
Niphargus plurispinosus sp. n., Crustacea, Amphipoda, Carpathian region, Slovakia, morphology, variability, phenology, ecology
There are over 300 species and subspecies described in the genus Niphargus Schiödte, 1849, distributed mainly in ground waters of Europe (
The situation in Slovakia is no different. Eight species of the genus Niphargus are currently known from Slovakia (
Three years of intensive study of superficial subterranean habitats (sensu
The aim of this paper is to present detailed morphology of this species and to provide some data on its autoecology.
All material was collected at the same locality: South-east Slovakia, south-east foothill of the Zemplínske vrchy Mts. (different to nearby Zemplén Mts. in Hungary), in Hatfa in the village Viničky (48°25'11"N, 21°44'59"E). The mountain chain Zemplínske vrchy Mts. is a small part of the Carpathians (covering an area of about 6 × 15 km, with the highest top at 465, 3 m.a.s.l.). The complicated geology includes Paleozoic (non-carbonate), Mesozoic (carbonate) and Tertiary (Sarmatian volcanism, sediments) deposits (
The locality Hatfa near the village Viničky is situated on the boundary of areas with different history and bedrock (limestone and volcanic pyroclastics), on the south-east foothills of the hills, close to the limestone area. The type locality is a small, permanent spring close to a private reservoir of groundwater (164 m.a.s.l.), with artificial drainage to the meadow at the east foothill (Fig. 1). Water temperature in the spring is relatively high all year round (10–13°C). Chemical parameters were pH = 7.08, conductivity = 1310 µS, concentration of Ca ions 18.6 mg.L-1 and concentration of Mg ions 32.1 mg.L-1. The stream bed varies from 5–10 cm in width and 1-3 cm in depth. All specimens were collected from a drainage ditch under the stones, leaf litter and grass using a tea-strainer (leg. A. Mock).

Locality (Viničky-Hatfa) of Niphargus plurispinosus sp. n. in Slovakia: A general view B detail of location where the specimens were collected (Photo: A. Mock).
Most specimens were preserved in 75% ethanol, with a few samples preserved in 96% ethanol for future DNA-analysis. One to three specimens from numerous samples were morphologically studied.
From all samples we took the smallest and the largest specimens. Adult females were distinguished by the presence of eggs whereas adult males were identified on the basis of having elongated exopodites of uropods III. All other stages were assumed to be correlated with body size (see in Variability chapter).
Specimens were boiled in KOH and chlorophenol; later they were dissected and mounted on slides using SWANN-medium. Fine details were examined using a Leica microscope with magnifications 100–400× using black field or phase contrast. All pen-drawing were made using camera lucida. Digital photos were taken with an Olympus DP10 camera mounted on a Leica microscope or stereomicroscope.
Symbols and abbreviation used in the text:
(⇚) very important taxonomic character;
(←) useful supporting taxonomic character.
All other abbreviations of morphological terms used in the text correspond with
http://zoobank.org/9900B10D-7230-44DD-86CB-F904F45EF08A
http://species-id.net/wiki/Niphargus_plurispinosus
Figs 2–9The species name was derived from the Latin words: plus, pluris (= more) spina (=spine, thorn); Niphargus with more then 1 dorsal spine (thorns) on the telson.
The type series was collected in the locality Viničky - Hatfa.
Holotype. Viničky - Hatfa 11 May 2013: one adult male (16 mm) in vial preserved in ethanol, (NHMUK 2014.381 Natural History Museum, London, Great Britain).
Allotype. Viničky - Hatfa 11 May 2013: one adult female with eggs (14 mm) in vial preserved in ethanol (NHMUK 2014.382NHM, London).
Paratypes. Viničky - Hatfa 11 May 2013: one large male (17 mm) in vial preserved in ethanol (NHMUK 2014.383 NHM, London); 1 dissected large male (20 mm) mounted in Swann-medium (NHMUK 2014.384 NHM, London); 1 dissected largest female (17 mm) without eggs mounted in Swann-medium (NHMUK 2014.385 NHM, London) and 1 dissected large male (20 mm) (in the authors collection).
Paratype serie. Viničky - Hatfa 19 June 2012 50 specimens of different stages in one vial preserved in ethanol (NHMUK 2014.386-395 NHM, London).
Paratypes. More than a dozen specimens deposited in Ljubljana, Slovenia (Collection of C. Fišer), the remaining samples in the authors collection.
2011: 26 May: 5 specimens (3 adult ♂, 2 adult ♀ with eggs); 14 October: 30 specimens (25 subadult ♀ and 5 subadult ♂); 23 November: 6 specimens (4 subadult ♀ and 2 subadult ♂);
2012: 12 January: 10 specimens (6 subadult ♀ and 4 ♂); 23 February: 32 specimens (22 juveniles, 4 subadult ♀ 6 subadult ♂); 21 March: 35 specimens (2 adult ♀ with eggs, 33 juveniles); 3 May: 19 specimens (1 adult ♀ with embryos, 1 adult ♂, 17 juveniles); 19 June: 17 specimens (1 subadult ♀, 1 subadult ♂ (both about 12 mm long), 10 juveniles (up to 8 mm), 5 neonates (not longer than 4 mm); 9 July: 16 specimens (15 juveniles and 1 neonate); 28 September: 10 specimens (all juveniles); 22 November: 33 specimens (32 juveniles, 1 subadult ♀);
2013: 11 May: 21 specimens (3 ♀ with embryos, 7♀ larger but without embryos (post adults), 7 ♂ more than 20 mm long (post adults), 25 juveniles), preserved in clear ethanol; part of material is in Ljubljana in C. Fišer’s collection.
All remaining material is retained in the authors´ collection.
Niphargus plurispinosus: Middle-sized, oblong species with small gnathopods, sexually dimorphic uropod III and sexually non-dimorphic uropod I in juveniles, but different in adults, and extremely different in postreproductive stage. Coxal plates as follows a) cx-1 rhomboid; b) cx-7 reduced, trapezoid-oval plate with elongate posterior corner and one seta close to posterior corner; other coxal plates (cx-2 to cx-6) without species-specific features. Epimeral plates - first two vaulted in posterior-ventral corner; 3rd angular (juveniles and adult males). Telson: lobes slightly narrowing distally, with even end, deep cleft: 70–80%; 2-3(4) dorsal spines arranged in a transversal row; c) terminal spines do not exceed 35–40% of telson length, and decrease with age down to 20%. Pereopods V-VII bases - elongate-oval, length ratios = 1.0/1.2/1.25; dactyli with long nail (30–40% of dactylus length) and one tiny spiniform thorn near nail base. Mouthparts: maxillae I inner lobe with 3 setae: 2 terminal and 1 subterminal; maxillipeds inner lobe with 3 lancet-like stout setae.
If it is not specified the characters are the same for both sexes.
Body shape. Longish and narrow (Fig. 2: 1). Body length: up to 20 mm (male), 17 mm (female). Colour: white-yellowish (living specimens); shortly after preservation in alcohol the body turns white.
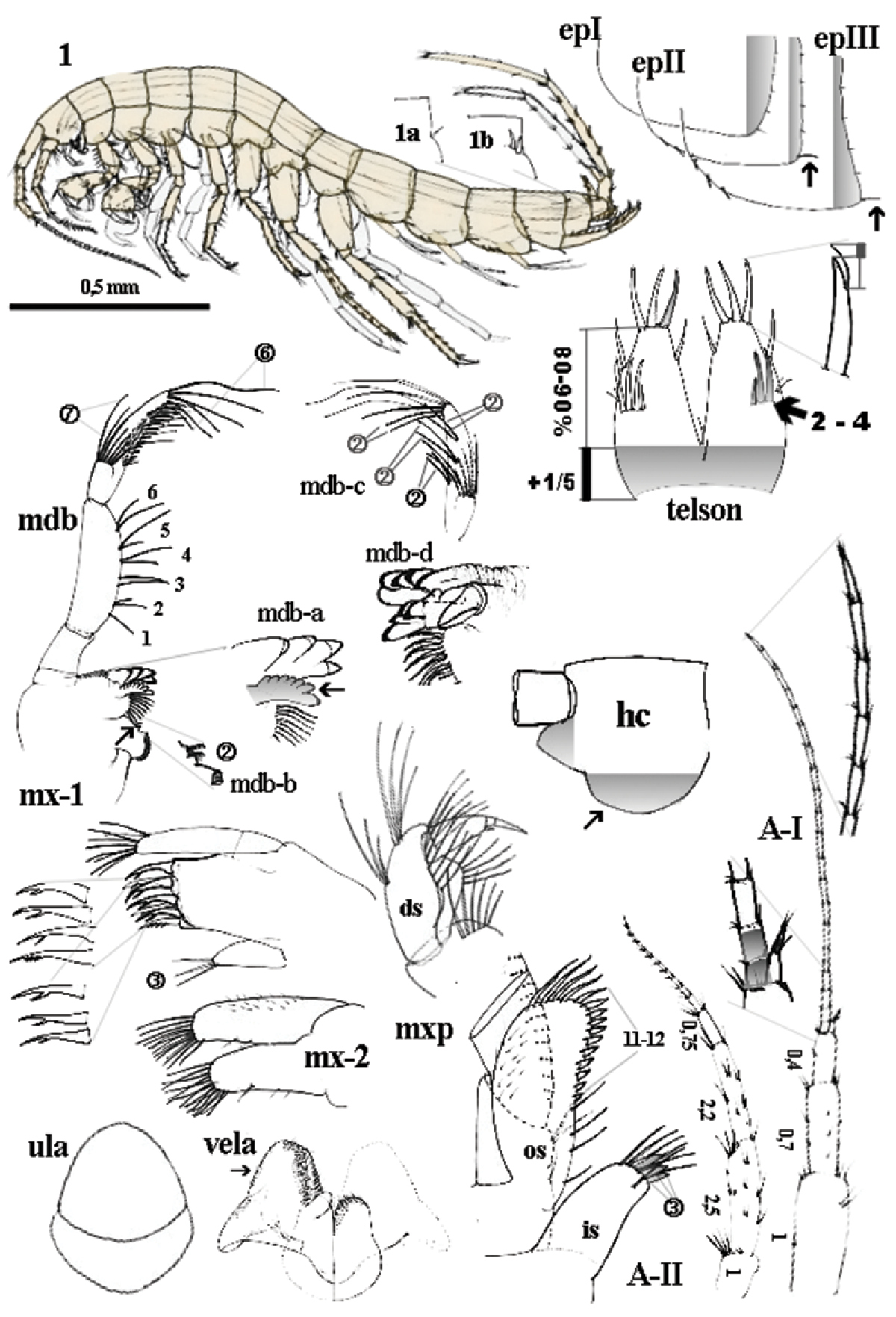
Niphargus plurispinosus sp. n.: 1 male, general view; 1a-1b dorso-later thorns; mdb - mandibula and details of mdb-a left incisor and lacina mobilis; mdb-b) two setae between bisserated thorns; mdb-c setae pattern on distal segment of mdb-palp; mdb-d, right incisor and lacina mobilis; mx-1 1st maxilla; mx-2 2nd maxilla; ula upper lip; vela ventral labium; mxp maxilliped: in inner segment os outer segment; ds distal segment of palp; epI-epIII epimeral plate I-III; A-I 1st antenna; A-II antenna; hc head capsula, left lateral view; telson, dorsal view. Not scaled, except of the general view of the male.
Head (Fig. 2: 1; hc). Short angular, without rostrum; anterior margin deep sinusoid (deep incision at the joint of AI and long, lobe); ventral (cheek) margin expressive vaulted. Surface of head capsule is smooth. Minute yellow spot on each side of head between AI and AII present in living specimens, but these quickly fade on preservation.
Antennae: 1st antenna (Fig. 2: A–I). length 35–40 % of BL; ratio between the three pedunclar segments is 1 : 0.7 : 0.4; flagellum with 20–21 segments of different sizes expressed as ratio to 1st peduncular segment: 0.17 (1st -2nd) - 0.18 (3rd-4th) - 0.10 (5th- 6th) - 0.20 (up to 21st); each segment with a few minute sensillae and one elongate bi-segmented aesthetasc along distal margin; accessory flagellum bi-articulated and short (shorter than 1st two segments of flagellum) with 4 sensillae on tip. 2nd antenna (Fig. 2: A-II): length 50 % of A1; peduncule segments ratio =1 : 2.5 : 2.2. Flagellum bears basal longer segment (0.7 of 1st segment) and <10 short segments of moderate size to 1st segment: 0.5 (1st -2nd) - 0.38 (3rd–4th) - 0.25 (up to 10th).
Upper lip (labrum) (Fig. 2: ula; Fig. 3: 1). Sclerotised, oval-quadrangular, 2-articulated: basal segment narrow, distal segment sub-oval; whole surface is smooth or with long submarginal suture in distal portion.
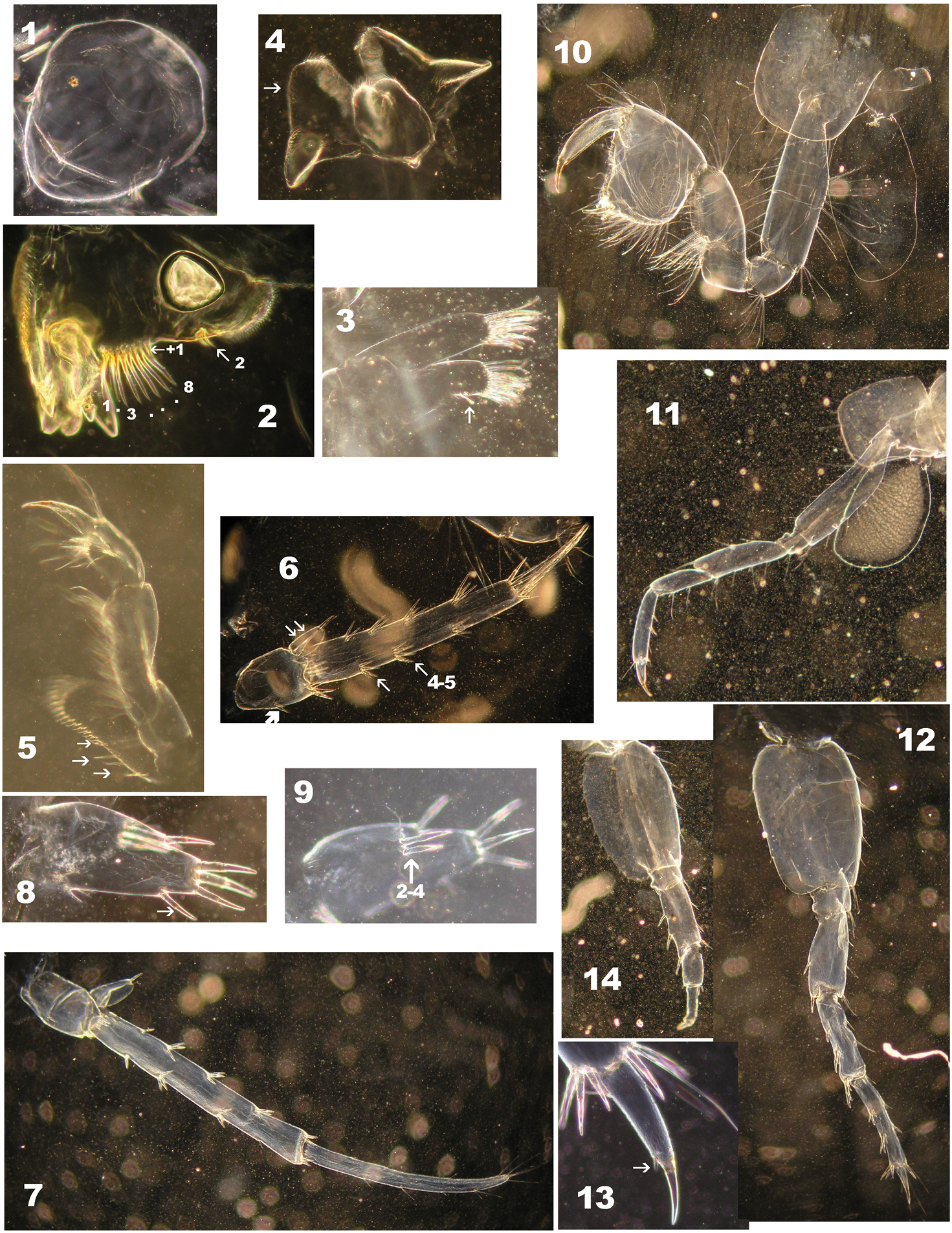
Niphargus plurispinosus sp. n.: 1 upper lip 2 right mandible 3 second maxilla 4 labium 5 half maxilliped (without inner portion) 6 3rd uropod (female) 7 3rd uropod (juvenile male) 8–9 telson (one lobe of male) 10 2nd gnathopod 11 4nd pereopod 12 7th pereopod 13 distal segment and 6th pereopod 14 6th pereopod: regeneration of segments behind basis (Photo: I. Hudec). Not drawn to scale.
Mandibles (Fig. 2: mdb). Right mandible (Fig. 2: mdb-a): incisor with 4 sclerotised teeth, lacina mobilis with 7 small, vaulted teeth arranged in short fan; the first one is larger (Fig. 2: mdb-a ←). Left mandible (Fig. 2: mdb-d): incisor with 5 sclerotised teeth; lacina mobilis with 4 sclerotised teeth. Both mandibles: between lacinia and molar a row of 8 longer and 1 shorter of thick, bi-serrated setae; 2 minute setae on middle position of the outer molar margin (Fig. 2: mdb-b; Fig. 3: 2 ←); molar plate sclerotised and one longer seta at the inner base of molar. Mandibular palp three segmented: the shortest basal segment without setae; the two distal articles are of equal lengths; middle segment with 6 transverse bunches of setae (each with 2 setae) along inner margin; distal segment with 1 (basal) transversal row of 7 setae (A-setae sensu
Maxillae I (Fig. 2: mx-1). Palpus 2-segmented, distal segment asymmetric vaulted with 14 longer terminal setae and 1 smaller seta on outer margin (near the tip); outer lobe 7 denticulated spines arranged in two rows (inner row 4 - outer row 3 teeth); spines with secondary structures (denticles) as follows: 6 uni-, 1 two-denticled; inner lobe with 3 setae (2 on tip and third one in subterminal position) (Fig. 4: 5).
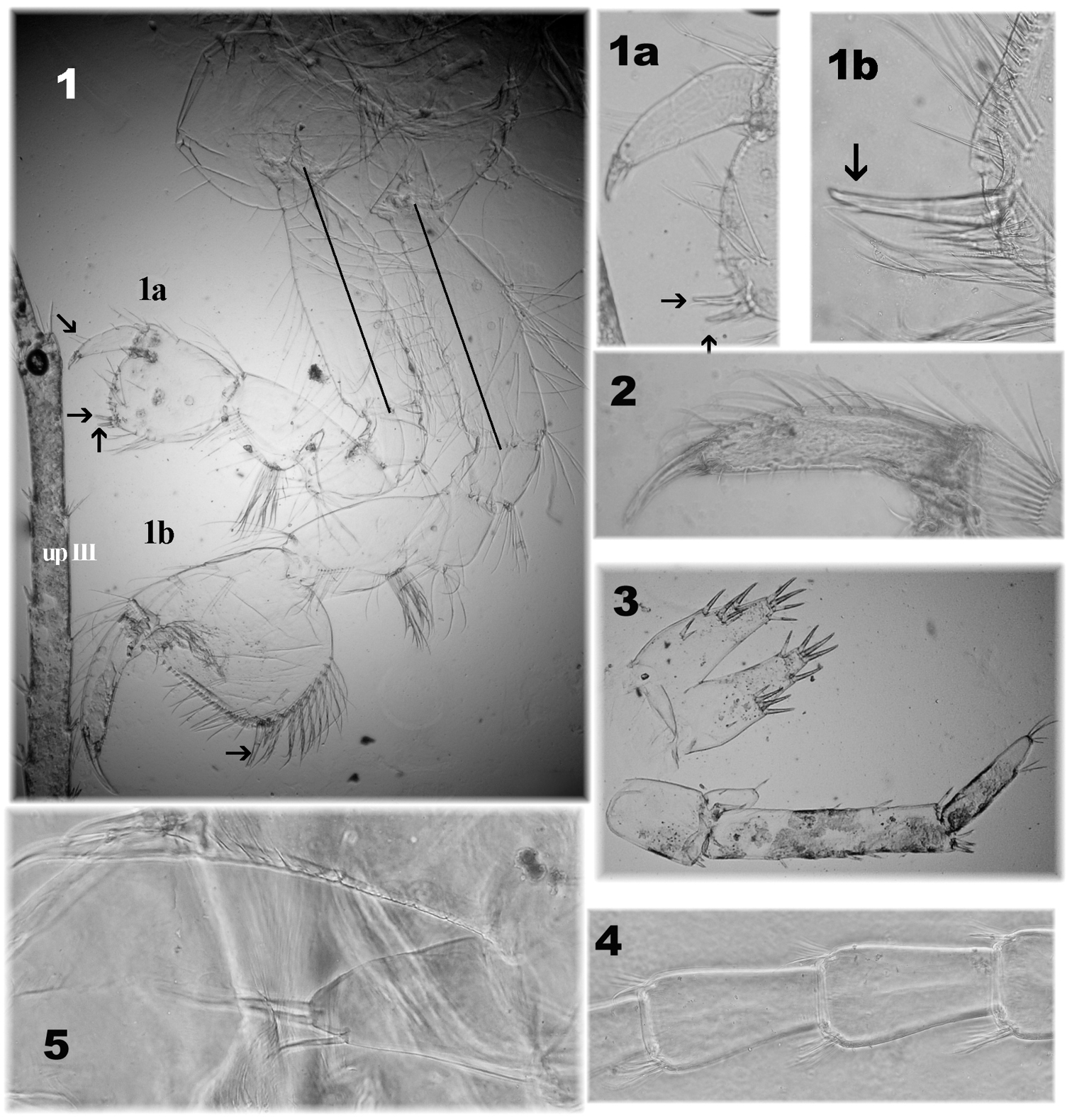
Niphargus plurispinosus sp. n. – “postreproductive” male: 1 gnathopods with detail of deformed gpI (1a) and normal developed gpII (1a) 2 gpII -setae on dactylus 3 telson and regenerated (?) upIII 4 segment setae on flagellum of AI 5 maxillae I: position of three setae on inner portion (Photo: I. Hudec). Not drawn to scale.
Maxillae II (Fig. 2: mx-2; Fig. 3: 3). Both lobes sub-equal in size, with apical setae (inner and outer lobe) and one seta in ventral margin of ventral lobe. Dorsal margins of both lobes with fine hairs.
Labium (Fig. 2: vela; Fig. 3: 4). Larger inner lobes trapezoid-like with sub-triangular flat posterior protruding on each portion; its outer distal (←) and anterior-inner carries fine feather-like setae. Smaller outer lobes compact and of subovoid shape are fine serrated on distal portion.
Maxillipeds (Fig. 2: mxp; Fig. 3: 5). Inner lobe short with 7 long setae, (6 marginal- plus 1 submarginal seta) and 3 flattened spine-like setae on apical part (←); outer lobe reach up to ½ of 2nd segment of maxilliped palp, with 4 isolated longer setae near base and crest of 9–11 flattened spine-like setae which increase distally along inner margin and are followed by 5 longer denticulated setae along distal arc. Outer surface finely setuled. Palp 4-segmented: 1st basal, subtriangular segment with one bunch of 4–5 setae on inner side; 2nd segment, (the longest one) with 14 transversally oriented rows of setae along inner margin and bunch of 7–10 setae close to distal end (outer side) and one bunch of setae near base of inner margin; 3rd segment small sub-oval with two long setae in the middle of dorsal margin and two rows of numerous setae around dactylus; 4th – dactylus with 1 short, bent denticle and 1 short spiniform seta on 2/3 length of ventral margin and one seta in 1/3 of dorsal margin; terminal nail about 1/3 of whole dactylus.
Coxal plates (Fig. 5: cx) flattened with isolated setae along ventral margin. CxI rhomboid-like, antero-ventral corner broadly subrounded; cxII–IV rectangular, angles rounded; anterior, ventral and posterior margins vaulted; cxV–VI similar with well developed anterior lobe ventrally with 4 setae along distal end (cxV) or 1–2 setae (cxVI); posterior part elongated posteriorly with two setae close to posterior ventral angle (cx V) or 4 setae along posterior margin (cxVI); cxVII reduced, trapezoid-oval plate with elongated posterior corner and one seta close to posterior end (←).
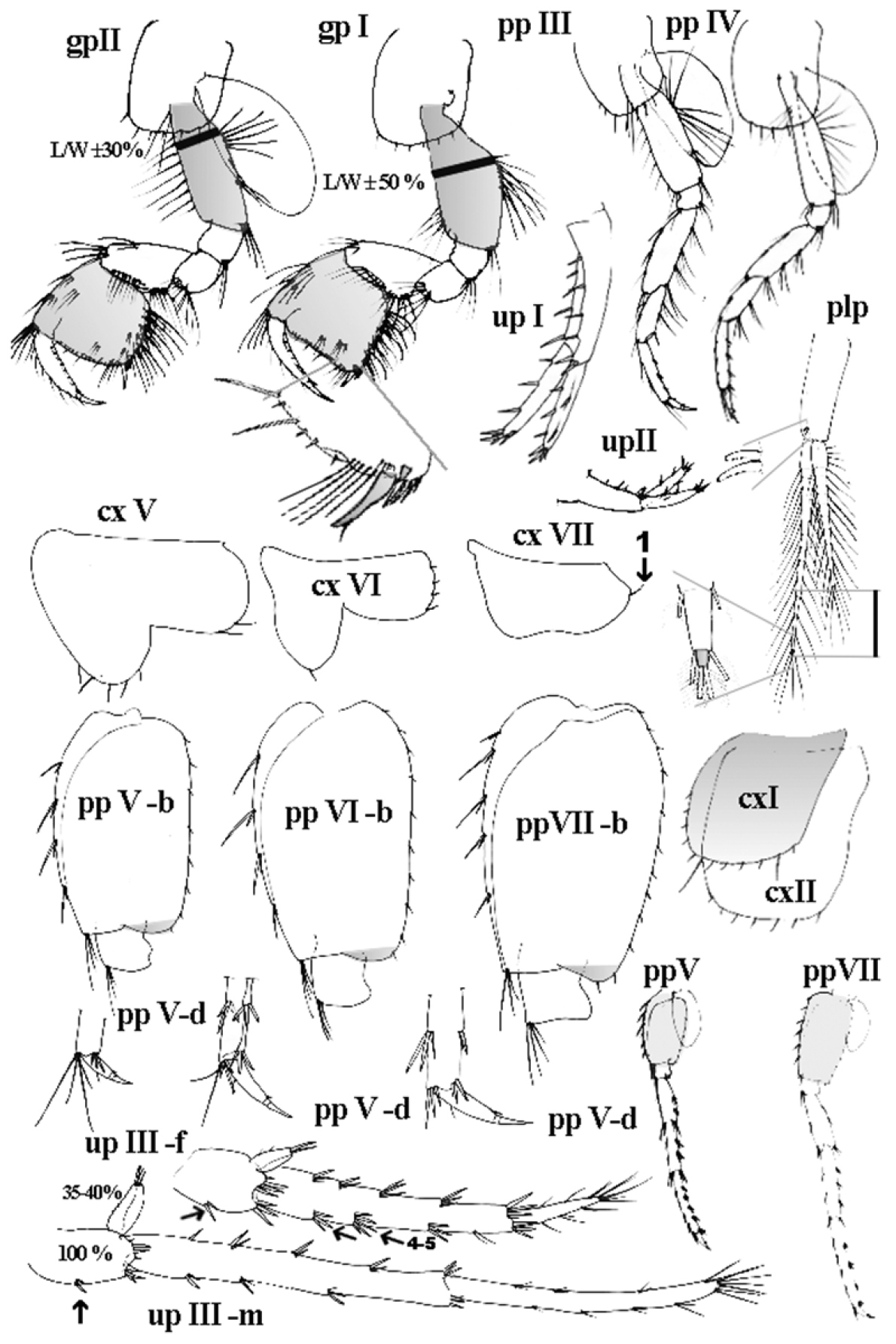
Niphargus plurispinosus sp. n.: gpI 1st gnathopod; gpII 2nd gnathopod; ppIII–ppIV, ppVI–ppVII 3rd to 7th pereopods; ppV-b – ppVII-b bases of 5th to 7th pp; ppV-d – ppVII-d distal part of 5th to 7th pp; plp 2nd pleopod; upI and up II 1st and 2nd uropod; upIII-f 3rd uropod of female; upIII-m 3rd uropod of male; cxI-cxVII 1st to 7th coxal plate. Remarks: shadow colour was used to emphasise of important character. Not drawn to scale.
Gills uniformly broad, asymmetric sub-oval, reaching from 4/5 (basis II) to 1/3 (basisVI) of basis.
Pereon appendages (Figs 3, 5, 6).
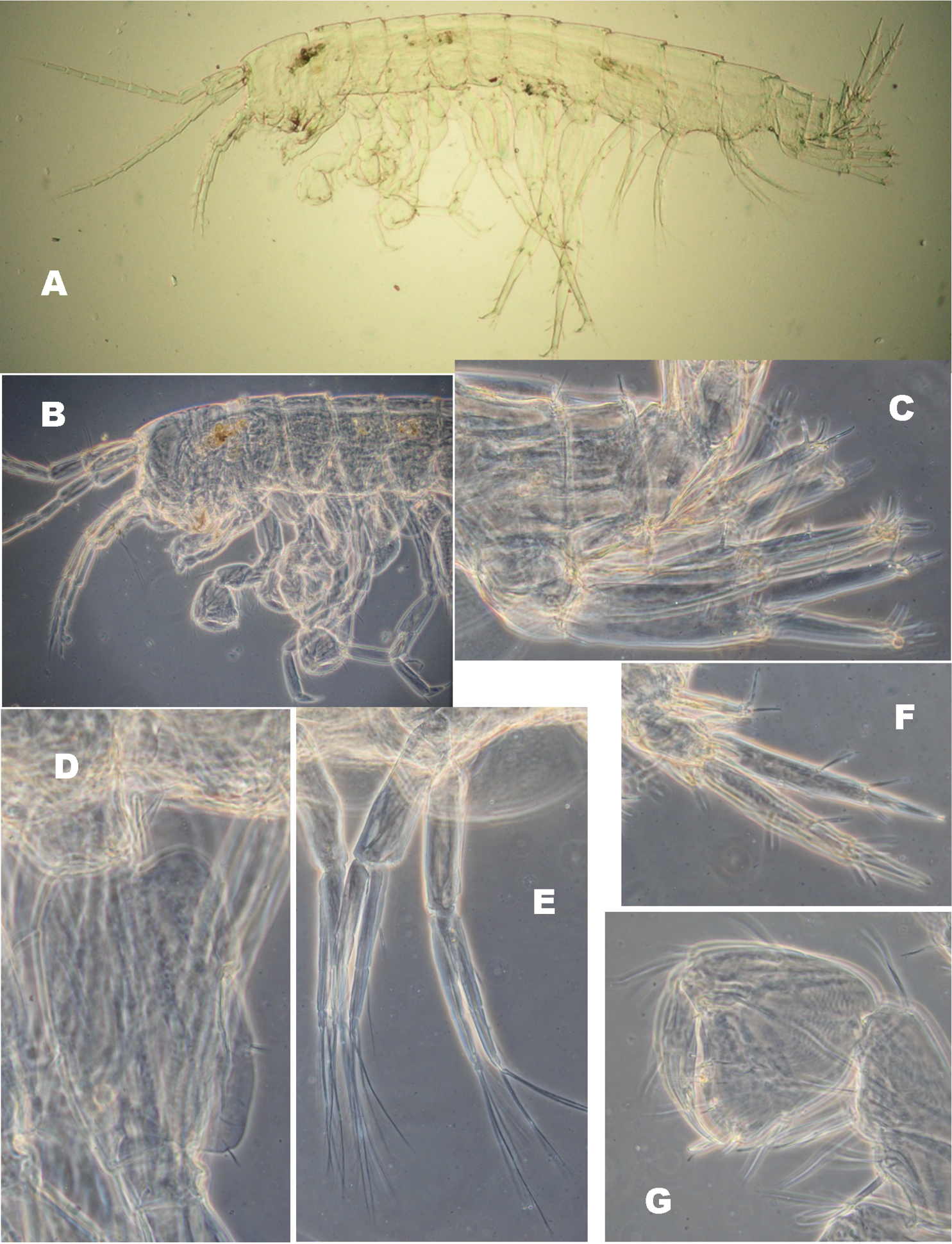
Niphargus plurispinosus sp. n. neonate: A general view B head and anterior part C 1st and 2nd uropods D basal segment of 7th pereopod E 2nd pleopod F 3rd uropod G 2nd gnathopod (Photo: I. Hudec). Not drawn to scale.
Gnathopods I (Fig. 5: gp-I; Fig. 6: G). Basis – trapezoid broad (L/W ±50%), laterally flattened; a group of longer setae close to anterior ventral angle; two groups of setae. Ischium – sub-quadrangular, bears 7–9 setae on posterior-distal angle. Merus – sub-angular (almost identical with ischium) bears transverse row of setae and a short row of submarginal setae near anterior-distal corner. Carpus – elongate sub-trapezoid with the longest dorsal margin; dorsal margin with 1 group of long setae almost on anterior-distal corner; ventral margin with expressive bulb in ventral base covered with expressive large setae on surface; a submarginal row of setae follows posterior margin; carpus length 60–65% of basis length and 110% of propodit length. Propodus – sub-quadrate; anterior margin with 3 transverse oriented rows of long setae (two rows along margin and third one almost on anterior-distal corner); distal margin (palm) is convex or almost even with 4 long setae each interrupted with 2–4 minute (thorn-like) setae. Palmar corner with the bunch of 4–5 longer setae close to strong palmar spine; one long, blunt pointed, thick seta; it is followed by 3 shorter, stronger and serrated, spiniform setae on outer side and one supporting minute stout spine on inner surface. Along posterior margin 6–7 transversal rows of numerous setae are present. Two groups of tiny, setae are present on outer surface (close to ventral corner). Dactylus – long (as maximal height of propodit); along anterior margin 6-7 longer, single setae (←); along inner margin a row of sparse minute setae.
Gnathopods II (Fig. 3: 10; Fig. 5: gp-II). Basis – trapezoid narrow (L/W ±30%), sub-oval in transsection; sparse row of long, sub-equal setae along anterior margin and three bunches of setae on posterior margin: a) numerous, long setae on basal angle; b) few setae almost in middle position; c) few setae near posterior-distal angle. Ischium – sub-quadrangular, bears 7–9 setae on posterior-distal angle. Merus – sub-angular (almost identical with ischium), bears transverse row of setae and a short row of submarginal setae near anterior-distal corner. Carpus – elongate sub-trapezoid, its dorsal margin the longest; dorsal margin with 2 groups of setae near anterior-distal corner; ventral margin with expressive bulge in ventral base covered with expressive large setae on surface; a submarginal row of setae follows posterior margin; carpus length 70–75% of basis and 120% of propodit. Propodit– subquadrate; anterior margin with 3 transversely oriented rows of long setae (two rows along margin and the third one almost on anterior-distal corner); distal margin (palm) convex with 3–4 longer setae each interrupted with two to four minute (thorn-like) setae. Palmar corner with the bunch of 4–5 longer setae close to base of strong palmar spine; one long, blunt pointed and thick seta and 3 stronger-spiniform serrated setae on palmar corner; one supporting minute stout spine on inner surface. Along posterior margin 6 –7 transversal rows of numerous setae are present. Four groups of doubled or triplet, tiny, spiniform setae are present on outer surface. Dactylus – long (as maximal height of propodit); along anterior margin 6–7 longer, single setae (←); along inner margin a row of sparse minute setae.
Propodits of both gnathopods sub-equal in size, the second one slightly larger. Compared to body size, gnathopods are small.
Pereopods III-IV (Fig. 3: 11; Fig. 5: ppIII, ppIV). Both subequal in morphology and size. Secondary spines of each segment have unknown taxonomic value. Distal ends of each propodus with 2 expressive long and 4 shorter seta-like thorns on anterior corner; 2 shorter seta-like thorns and 2 stout thorns on ventral corner correspond with pereopods V. Dactyli III-IV each with long nails (up to 30-40% of dactylus length) with dorsal plumose seta in the proximal third of anterior margin of the article, and one tiny spiniform thorn near the nail base. Spiniform spine is slightly bent to dactylus.
Pereopods V-VII (Fig. 3: 12–13; Fig. 5: ppV – ppVII). Sub-equal in morphology but different in length. Ratio of pereopod V-VII length = 1.0/1.4/1.5, where length of 5th pereopod is almost equal to that one of ppIII and ppIV. Bases V–VII elongate-oval (←), with convex anterior margins and almost straight posterior margins (←), all almost without ventro-distal lobes; length/width ratio = 1.00/0.55–0.65; L-ratio of bases = 1.0/1.2/1.25. Along anterior margins 4–5 slender spiniform setae and one bunch of setae-like thorns on antero-ventral corner; along posterior margins 8–11 small setae. Distal ends of propodit V–VII with characteristic combination of long setae-like thorns and stout thorns on each pereopod: V- equal to pereopods III-IV; 6th VI-with 2 slender setae-like thorns and 2–3 stout thorns on anterior angle and 2 stout thorns on ventral corner; 7th with 2 slender setae-like thorns and 2–3 stout thorns on anterior angle and 2 stout thorns on ventral corner. Morphology and setal patterns of dactyli V-VII are identical to those in dactyli III-IV, however the length of each nail can be variable, probably it is the result of their break or mechanical wear out.
Pleosome section (Figs 2, 5)
Pleonites I–III (Fig. 2: epI-epIII). Each composes from two different parts: dorsal and ventral part with epimeral plates on each side. Dorsal part of all pleonites with distinct anterior margin (minute hump on anterior part of distal angle); 6-8 fine setae along dorsal margin (under cover glass it is a part close to the posterior-dorsal corner). Sub-rounded, ventral epimeral plates are clearly distinguished from dorsal part. Epimeral plate I (epI) anterior-ventral corner narrow vaulted forms blunt angle with ventral margin; ventral margin convex, broadly vaulted (without thorns); posterior-ventral corner convex and broadly vaulted. Along posterior margin 5-7 setae (the first one is the longest). Epimeral plate II (epII): anterior-ventral corner broadly vaulted; ventral margin slightly convex with 2 submarginal stout thorns; posterior-ventral corner broadly vaulted. Along posterior margin 5-7 seta-like thorns (the fist one is stouter and the longest). Epimeral plate III (epIII): anterior-ventral corner broadly vaulted; ventral margin slightly convex with 3 submarginal thorns; posterior-ventral corner angular or perpendicular (←) with blunt tip. Along posterior margin 5-7 seta-like thorns (the first one is slightly longer).
Pleopods I-III (Fig. 5: plp). Uniform: each with smooth tubular protopod and two retinacules on distal end; two rami (longer one with 13 articles; shorter one with 15 articles). Each segment bilaterally setuled on distal end, except for the proximal segment. First basal segments 3 to 4-times longer than next segment with row of 4 to 5-times shorter setae on outer margin on shorter arm and smooth on longer segment. The most distal segment is minute and conic.
Urosome section (Figs 2, 3, 5, 7, 8).

Niphargus plurispinosus sp. n. – male: ontogenetic transformation of upI (6–8), upIII (1–5) with detail of endopodite (3–1, 4a1, 5–1) and presence of black-brownish callus (▼) after extraction and wounding of extremities, or after a bite (4–2) 1, 6 neonate 2–3 juvenile 4, 7 adult 5, 8 postreproductive male 9 gpII of neonate (Photo: I. Hudec). Not drawn to scale.
Urosomite I (Fig. 2: 1-1a, 1b) posterior-dorsal corner with 1 weak submarginal seta; ventrally 1 short, slender thorn near insertion of uropod I. Urosomite II posterior-dorsal corner with 2 submarginal spiniform thorns (one subtile and one stouter). Urosomite III without setae.
Uropods I-III: UpI and upII are morphologically similar but the first one is almost twice as long as upII; upIII is sexually dimorphic.
Uropod I (Fig. 5: upI; Fig. 7: 7). Protopodit without flap on its ventro-distal end; it is longer to both, subequally long distal rami (±1.1–1.2) protopodite bears 9 thick dorso-lateral spines, arranged in two rows (5+4). The endopodite is longer and rod-like. However with age the exopodite is gradually enlarged and transformed into a club-like structure (Fig. 4); both bear spines arranged in two rows; longer flexibile setae are on distal half; 5 thick spines of different size (two longer) on distal end. Uropod II (Fig. 5: upII): length of endopodite is 1.05–1.15 of length of exopodite and both are shorter to basipodite.
Uropod III - male (Fig. 5: upIII-m; Fig. 7: 4). Up to 35-40% of body length (all following measures are valid for adults). Base sub-oval, short (= 1/4 L of basal segment of exopodite) with numerous, grouped spines along distal margin (around base of exopodite) and 2 stout spines in the middle of ventral margin (←). Short endopodite (up to 35-40% of base L (←)) bears 2–3 short thorns on distal end and 1–2 minute spines on outer lateral margin. Two-segmented exopodite rod-shaped; basal segment slightly shorter (90-95%) to distal segment; basal segment with 4 groups of spines along ventral margin and 5 groups of spines along dorsal margin; distal segment with 4 groups of longer setae along ventral margin and 3 tiny setae along dorsal margin; distal portion with 4–6 clusters of long setae.
Uropod III - female (Fig. 3, Fig. 5: upIII-f, Fig. 8) robust, shorter then in male; base suboval (1/4 of basal exopodite segment) with two spines on ventral margin (←). Moderate short endopodite (up to 45% of base length) bears 2–3 short thorns on distal end and 1-2 minute spines on outer lateral margin. Two-segmented exopodite conical, narrowing distally. Basal segment 3-times longer than distal segment. Basal segment with 9 groups of spines (5 along dorsal- and 4 along ventral margin). Other group of thin longer spines on ventral margin (close to distal end). Distal segment only with groups of longer setae-like spines.
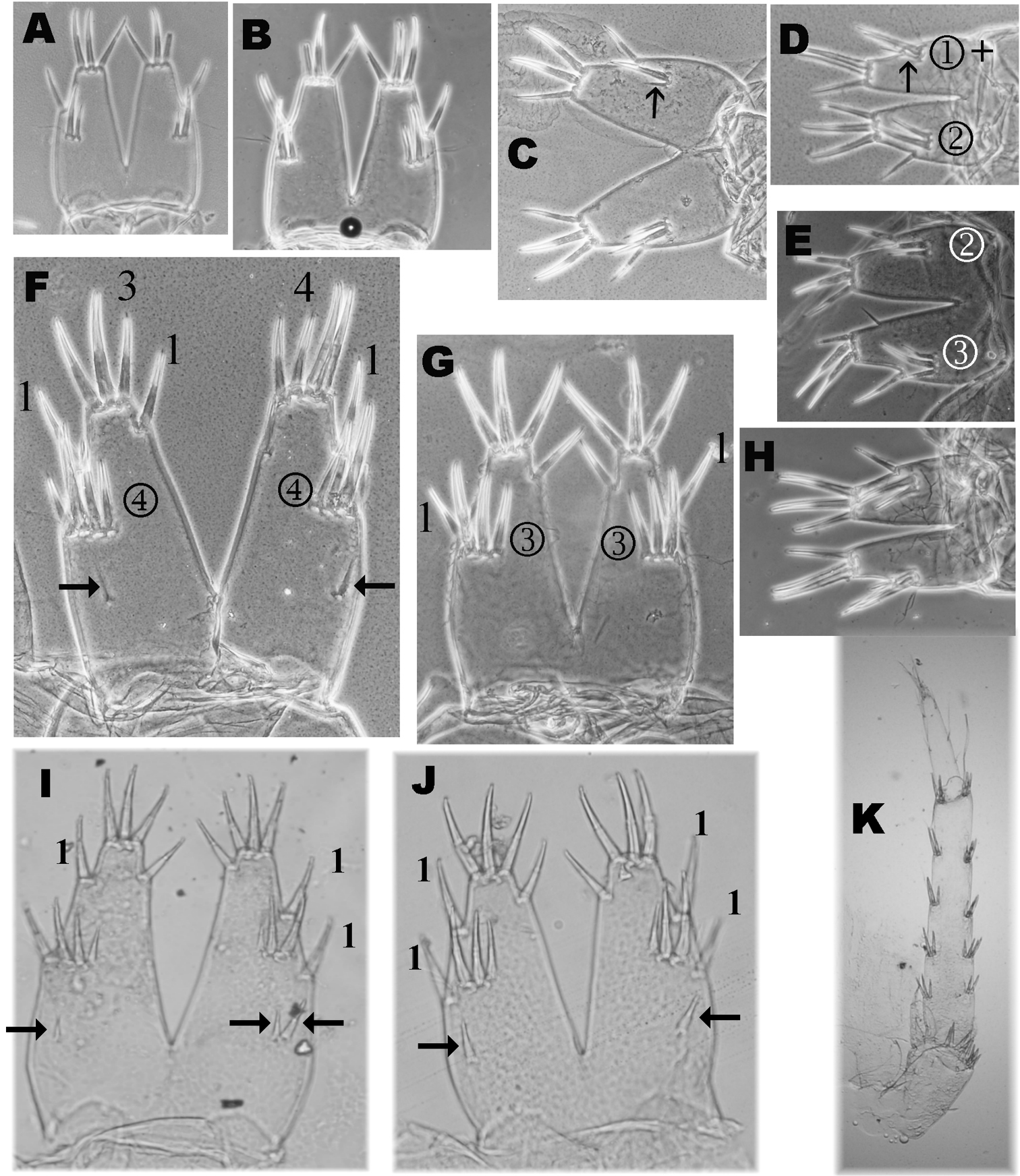
Variability of telson setae of Niphargus plurispinosus sp. n.: A–E, H juveniles (males and females) F adult male G adult female I postreproductive male J postreproductive female K 3rd uropod of female (Photo: I. Hudec). Not scaled.
Telson - both sexes (Fig. 2: telson, Fig. 3: 8–9; Fig. 8: A–G) angular, length/width = 1.1–1.2, with deep cleft 80–90% of telson length (⇚); lobes narrowing distally, almost even in terminal part. Apical telson spines relatively short (35–40% TL) (⇚) where distal lateral flagellum protruding over the terminal end of spine. Spine position (per lobe): 3-4 terminal (apical) spines; 1(2) outer lateral spine and 1 inner lateral (mesial) spines in ¼ TL (from distal end). 2–3(4) dorsal spines are arranged in one transverse row (⇚), situated more-less in ½ TL; however the first (outer) dorsal spine is probably outer lateral spine (Fig. 8: F). Pair of plumose setae inserted mid-laterally. 2–4 thin, relative long, spines just below cleft. One slender, supporting dorsal spine which can be found before dorsal thorns was recoded in the one largest male (Fig. 8: F ←).
The main problem when identifying Niphargus plurispinosus (and probably in all species of the Niphargus genus) is to distinguish principal characters sensu International Code of Zoological Nomenclature (
a) Ontogenesis and heterochrony. We identified expressive differences in external morphology among neonates (L up to 4 mm), first developing stage (L up to 6 mm), subadults (about 12 mm by both sexes), adults (L < 15 mm in females, 16 mm in males) and the absolutely largest specimens (L >17 mm in females, 20 mm in males). Without knowledge of the comparative morphology at least of first stages, adults (described above) and the largest (postreproductive?) specimens of Niphargus plurispinosus it is quite easy to identify them as 2–3 separate co-existing subspecies.
Basic difference for the neonate and (or) the first stages of Niphargus plurispinosus are:
1. Head is large relative to BL (Fig. 6: A–B). It bears shorter A1 (25–30 % of BL - because of lower number (8–10) of flagellar segments); A2 reach up to 60 % of A1 and bears 4–5 longer segments in flagellum. 2. Gnathopods: both gnathopods with modified subquadrangular propodits with 2-3 transverse rows of few setae along ventral margin (Fig. 6: G); number of transverse rows increase with the individual size of specimens (both as the number of setae in rows). Dactylus with only 2-3 setae along outer margin. (⇚) 3. Pereopods V-VII: Bases V–VII oval with expressive ventro-distal lobes, with 2 slender setae along anterior margin and 4-5 relative long setae along posterior margin (Fig. 6: D). Size of all three bases elongated but ventro-posterior lobe decrease with age; setae along anterior and posterior margins increase in number with size. 4. Telson bears only 3 apical spines, 2 dorsal spines, and one outer lateral spine more-less in mediate distance between apical- and dorsal spines on one lobe (Fig. 8: A-B). 5. Pleopods I -III: protopods without retinacules and 4-5 elongate articles in both rami (Fig. 6E). 6. Uropod I: exopod and endopod are equal or subeqaul in size; both with small number of spine-like setae (Fig. 6: C; Fig. 7: 6). 6. Uropod III: no sexual dimorphism at this size; (⇚) all specimens resemble females with reduced number of spines (Fig. 6: F; Fig. 7: 1). The sexual dimorphism is expressed in later (juveniles stages and preadults) as a continuous elongation of distal segment of exopodite (Fig. 7: 2-3) up to same length to basal segment by adults (Fig. 7: 4).
Similar patterns of postembryonic differentiation were noted also in Niphargus aggtelekiensis and Niphargus tatrensis (
The largests males (over 20 mm) and the largests females (17 mm), bear extreme cases of setation. Some character may change of in the following characters:
1. First antenna comparatively shorter to BL (up to 30 %); flagellum with <23 segments each segment with few minute sensilla but without bi-segmented aesthetascs (Fig. 4: 4). 2. Gnathopods (Fig. 4: 2): dactylus bears up to 12 single and doubled long setae (⇚) along outer margin; 3. Uropod I (Fig. 7: 8): exopod and endopod are extremely different in size (⇚); endopodite is markedly long (more than 2-times longer than exopodite) of club-like shape with reduced number of thick setae and multiplying longer setae which are restricted to the distal half of endopodite. This type of uropod resemble Niphargus cf. stygius (Schiodte, 1847) (
Specimens in probably postreproductive stage were recorded only once (11 May 2013).
b) Individual condition. The number of setae on all appendages changes with ontogenesis (
c) Regeneration or degeneration. We found that damaged extremities and body parts form blackish calluses within 12 hours of being damaged. The calluses persist after preservation and their colour is stable after the lightening process in different media (Fig. 7: 4-2, 7▼). An expressive case of regeneration was observed in a single specimen (subadult male) on the 5th pereopod (Fig. 3: 14): base normally developed, but the rest of the limb was extremely reduced. Specimens with different number of segments on AI- and AII-flagellum and males with different upIII might be probably considered as different stages of regeneration.
On the other hand the largest males (over 20 mm) were commonly found with different asymmetric gnathopods (Fig. 4:1, 1a, 1b, 2), different up III (Fig. 4: 3) damaged AI- and AII-flagellum (reduced number of segments very often without aesthetascs on AI, Fig 8: 4). However we are not sure if these damages are caused by attack, or age, or any kind of degeneration. Predatory behaviour and cannibalism were already observed in some niphargid amphipods (
The abundance (=number of animals per visit, when we collected all specimens) of Niphargus plurispinosus in the surface water near the spring ranged from 6 to 68 specimens in 2012 (227 ex. for the whole year). However, less than 8 % of all specimens were adults (12 males, 2 females with eggs and one female with embryos). Ovigerous females were recorded only in spring (March - May). Neonates were reported in June (15 inds.) and July (1 ind.). Juveniles and subadults prevailed in all samples and they were recorded throughout the year (Fig. 9: A). It looks as they leave subsurface actively to search for food in surface water. They were permanently found in the surface water throughout the year without sign of disfunction in mobility or presence of dead specimens. The only exception was in May 2013 of accumulated dead and dying animals. This may have resulted from water fauna poisoning when the owner of the reservoir disinfected the drinkable water by using chemicals with chlorine. Such events seriously endangered the stygophile fauna (
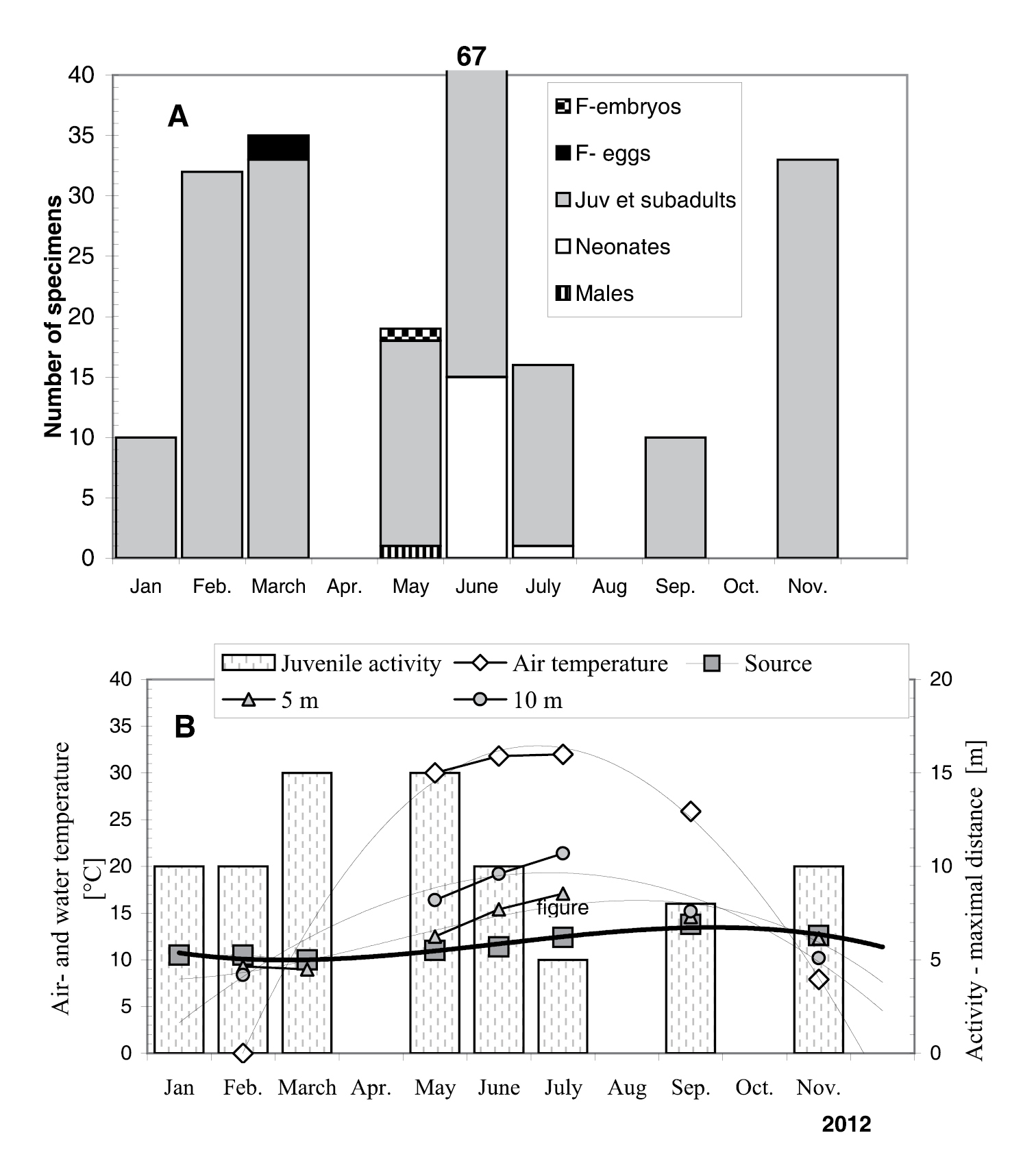
Relative abundance (A) and moving activity (B) of Niphargus plurispinosus sp. n. in relation to temperature gradient in the artificial channel of type locality in 2012. Air and water temperatures were measured during collecting of amphipods (around 2.00pm). Notes: F-eggs = females with eggs, F-embryos = females with embryos. Source, 5 m, 10 m = water temperature at various distances from the spring (source).
Light-yellow eye spots were seen in a few living adults, but we found no rudimentary eyes. These spots were smaller than those in Synurella ambulans (Müller, 1846). The yellow pigment disappeared within 2 days of preservation in alcohol.
All specimens of Niphargus plurispinosus were found in the shallow ditch low flow volume, up to 15 m away from the source; depending on age. Only juveniles (up to 8 mm of length) were found at the maximum distance (adults up to 5 m; subadults up to 10 m). Water temperature of the spring varied from 10.5°C (January 2012) to 13.8°C (September 2012) (Fig. 9: B). The distribution of specimens along the channel correlates with the water temperate gradient along the channel (Fig. 9: B), suggesting that water temperature over 17°C (July) may be a limiting factor for juveniles. This is in agreement with the known biology of hypotelminorheic habitats (
Niphargus plurispinosus was the only amphipod species found in the locality. It was collected only from a narrow (width = ± 10 cm) artificial drainage ditch, which originates from the small seep spring in the meadow (Fig. 2) and after 200 m it flows through the underground tube into a small stream with dense population of Gammarus balcanicus Schäferna, 1922. Such a semi-artificial locality can be considered as a unique natural laboratory for the study of biology of subterranean species that penetrate the surface.
The high number of dorsal spines on the telson lobes is typical for the new described species. But in the determination of the species from the genus Niphargus at least three additional characters need to be checked. In Niphargus plurispinosus these three additional characters are: small gnathopods, sexually dimorphic uropod III (adults and postreproductive stages) and sexually non-dimorphic uropod I in juveniles, but different in adults, and extremely different in postreproductive stage. For comparative purposes, we list three groups of species that share at least two out of the three traits mentioned above.
The first group consists of species with small subequal gnathopods and sexually dimorphic uropod III, but also sexually dimorphic uropod I. Several species sharing this combination have all or some of dactyls of pereopods III-VII armed with at least two spines (e.g.
The second group consists of species with sexually non-dimorphic uropod I, sexually dimorphic uropod III and unequal gnathopods, where the second propodus is large and its size largely exceeds the size of the first gnathopod: Niphargus stygius Schiödte, 1847 (
Finally, a species that share all three aforementioned traits is Niphargus sphagnicolus Rejic, 1958. The difference between Niphargus sphagnicolus and Niphargus plurispinosus includes the setal pattern on gnathopod dactyli I-II (single seta in Niphargus plurispinosus, several setae in groups in Niphargus sphagnicolus); number of spines at the base of uropod I (1-2 in Niphargus sphagnicolus, only one in Niphargus plurispinosus), lower number of submarginal ventral spines at epimeral plate III (3 in Niphargus plurispinosus, 4-5 in Niphargus sphagnicolus) and higher number of setae in maxilla I palpus (up to 9 in Niphargus sphagnicolus and 14 in Niphargus plurispinosus).
A detailed morphological description of Niphargus plurispinosus sp. n. based on large annual samples is presented. This is the first new species from Central Europe for several decades, showing that even this long-investigated territory has higher diversity potential than was supposed; similar situation is also in some other Carpathian regions (
There is a very low probability (less than 10%) that all stages will be obtained in one sample. It seems that for the description of a new Niphargus is necessary to obtain:
1. One abundant sample (dozens specimens of different size - including juveniles and adults); such abundant and heterogenous sample is almost impossible to obtain except spring, in such cases as studied in Niphargus plurispinosus.
2. One long series of samples (1 year) from one locality, collected monthly.
When describing new species it is very important beside the adult to describe also neonate and if possible also largest specimens in post-reproductive (senior) stages. The principal problem is expressive heterochrony (
This work was funded by the Slovak grant Agency (project VEGA 1/0139/09). We thank Cene Fišer (Ljubljana, Slovenia) for critical readings of the manuscript and for providing valuable comments and recommendations. Our thanks belong to both anonymous reviewers, Oana Moldovan (Cluj, Romania), as editor of the Subterranean Biology and Louise Maurice (Wallingford, UK) and Graham S. Proudlove (Manchester, UK) who considerably improved the manuscript by useful comments and linguistic revision, respectively.