Research Article |
|
Corresponding author: David C. Culver ( dculver@american.edu ) Academic editor: Oana Teodora Moldovan
© 2018 Heather Gilbert, Jenna Keany, David C. Culver.
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation:
Gilbert H, Keany J, Culver DC (2018) Response of shallow subterranean freshwater amphipods to habitat drying. Subterranean Biology 28: 15-28. https://doi.org/10.3897/subtbiol.28.30700
|
Abstract
The ability of three amphipods that occupy shallow subterranean habitats in the lower Potomac Basin of the US (hypotelminorheic), which often dry out seasonally, to withstand desiccation by burrowing in clay was investigated. Both Crangonyx shoemakeri, a wetland species, and Stygobromus tenuis, a subterranean species, burrowed in clay in the laboratory after surface water was removed. Gammarus minus, a spring species, did not. All three species exhibited behavioral changes as the habitat dried out.
Keywords
Adaptation to desiccation, burrowing, Crangonyx shoemakeri , Gammarus minus , Hypotelminorheic, shallow subterranean habitats, Stygobromus
Introduction
Freshwater habitats, such as wetlands and seasonal streams, typically first order streams, are subject to drying. Water level patterns, or hydroperiods, are an important way to categorize wetland habitats (
In contrast with wetland habitats, caves and other subterranean habitats are typically among the most environmentally stable freshwater ecosystems (
Burrowing to avoid desiccation is well known among wetland invertebrates, especially among species that have no resting stage and are unable to disperse, such as amphipods (
While there is little discussion of wetlands in the cave and karst literature (see
– A persistent wet spot that may dry up seasonally
– Underlain by clay or other impermeable layer
– Water exiting via seepage flow, typically from a slight depression
– Presence of blackened leaves
– A subterranean fauna, with species lacking or having reduced eyes and pigment
The hypotelminorheic is a subterranean habitat because it is aphotic except at its exit at a seep or seepage spring, where some light penetrates the leaf litter. We follow Williams’ (2017) distinction between seeps and springs. He defines seeps as comprising groundwater that emerges over a more diffuse area [than springs], with a rate of flow that is generally insufficient to form an outflow spring. The exit point, or seep, is an isolated wetland, rich in organic carbon from decaying leaves, and is in turn habitat for amphipods. This, in many ways unremarkable habitat, is made remarkable by its fauna. Of the 56 species in the exclusively subterranean amphipod genus Stygobromus found in the eastern United States 12 (21%) are found primarily in hypotelminorheic habitats (
With increased monitoring and sampling of seeps in the mid-Atlantic region of the U.S. (e.g.,
In the course of sampling seeps in parklands of Washington, DC, specifically in National Capital Parks-East (Piscataway, Oxon Cove, Anacostia, Fort Dupont, Shepherd Parkway, and others), it was found that the macro-crustacean fauna was dominated by two species of amphipods – Crangonyx shoemakeri and a Stygobromus species in the tenuis group – and the isopod Caecidotea kenki, (Keany et al. 2018). The Stygobromus species lacks eyes and pigment, and C. kenki and C. shoemakeri have reduced eyes and pigment. The undescribed species of Stygobromus is likely limited to the Anacostia River drainage in Washington, DC, and adjoining parts of Maryland. C. shoemakeri is a common inhabitant of wetlands in National Capital Parks-East and elsewhere in its range from southcentral Maryland to southcentral Virginia (
Our goal in this study was to determine whether the ability to burrow into clay substrates is an important factor in determining the presence of Stygobromus species and Crangonyx shoemakeri in seeps and the general absence of Gammarus minus. In order to investigate this, we conducted laboratory experiments monitoring the behavior and success of the three amphipods when water was systematically removed from a clay-filled petri dish.
Methods
Species
Three species were studied – Crangonyx shoemakeri (Hubricht and Mackin), Gammarus minus (Say), and Stygobromus tenuis potomacus (Holsinger). S. tenuis potomacus was used rather than the undescribed species from the tenuis group because there were not sufficient specimens of the undescribed species available. The two species are very similar morphologically and ecologically. All three species were found in seeps in the parklands of the District of Columbia, although G. minus was only found in seepage springs, typically with a permanent outflow, and usually during the Spring when water levels were at their highest.
Specimens used in this study were taken from seeps and springs in C & O Canal National Historic Park under permit CHOH-2016-SCI-0023. These individuals were obtained using standard collecting procedures as described by
Laboratory methods
The behavioral study was conducted in a Forma-lab Walk-in Environmental Room with a constant temperature of 5 °C. Glass pyrex dishes, with a volume of approximately 470 mL, (10 cm diameter and 4 cm depth) were lined with 30g of dry VWRTM clay, creating a clay layer approximately 2 cm deep. This clay was homogenized using a mortar and pestle and was sifted through a 125μm mesh sieve to ensure consistent grain size. Sixty mL of Deer ParkTM spring water were added to each of the glass pyrex dishes, creating a water depth of approximately 2 cm.
The three species of amphipods – S. tenuis potomacus, C. shoemakeri and G. minus – were studied separately. Within one week from the date of specimen collection, five individuals, all adults, of the same species were added to each dish. Ovigerous females were not used. After a 48 hour period of acclimation to the pyrex dishes, the trials began. This acclimation period was chosen based on the 2–10 day amphipod acclimation period recommended by
Six dishes were designated for each species; three served as the control while the other three were used as experimental dishes that simulated drying conditions. Each trial lasted five weeks.
A supplemental experiment was completed to determine the amount of water loss (retention) of the clay over the course of eight weeks. The dry weight of the clay was compared to the weight of the clay after one to eight weeks of drying. Measurements began at the end of week 1, after all surface water was removed.
During the course of the drying experiment, behavioral observations were made for 60 second intervals three times a week. The following behaviors were recorded if one or more individuals was observed with this behavior during the 60 second interval:
1 Walking (at least one body length)
2 Swimming (at least one body length)
3 Burrowing (direct observation or buried in clay)
4 Resting (resting for more than 10 seconds)
5 Interaction (with other amphipods)
6 Other (amplexus,etc.)
The relative frequency of these behaviors is not a direct measure of the time spent at each behavior because a behavior was given the same weight whether one or five amphipods was displaying the behavior. This indirect measure was necessary because it was impossible to track each individual separately. All observations were made under red light.
Data analysis
Both survival and behavioral data were analyzed used log-likelihood tests (G-tests), comparing observed and expected values in each category (e.g., walking in control dishes). G, which is distributed as χ2 was calculated as follows:
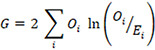 where Oi is the observed number of cases in category i and Ei is the expected number of cases in category i.
where Oi is the observed number of cases in category i and Ei is the expected number of cases in category i.
Results
Gravimetric measurement of moisture levels of the clay at the start of each trial averaged at 65.5% and reduced to an average of 54.2% after the first week of drying. Moisture levels then reduced to 34.4% at week two, and remained relatively consistent from week two to week nine, at around 30% (Fig.

Overall, there was a 25% increase in mortality for G. minus when subjected to drying (Table
The three species showed differences in behavior, both between experiment and control, but also between species (Fig.
Results of experiments on effect of drying. All runs began with 15 individuals.
| Species | Start of Trial | Control-surviving at 4 weeks | Experiment-surviving at 4 weeks | Difference | Log-likelihood | p |
|---|---|---|---|---|---|---|
| Gammarus minus | 7/20/2016 | 15 | 8 | -7.00 | 5.15 | <0.05 |
| 7/20/2016 | 15 | 7 | -8.00 | 4.44 | <0.05 | |
| 8/9/2016 | 15 | 10 | -5.00 | 3.45 | <0.10 | |
| 1/8/2017 | 2 | 2 | 0.00 | 0.00 | ns | |
| 1/8/2017 | 1 | 2 | 1 | 0.27 | ns | |
| Total | 48 | 29 | -19 | 7.61 | <0.010 | |
| Crangonyx shoemaker | 7/20/2016 | 13 | 12 | -1 | 0.53 | ns |
| 8/9/2016 | 15 | 15 | 0 | 0.00 | ns | |
| 1/8/2017 | 15 | 15 | 0 | 0.00 | ns | |
| 1/8/2017 | 15 | 15 | 0 | 0.00 | ns | |
| 1/8/2017 | 12 | 15 | 3 | 1.95 | ns | |
| Total | 70 | 72 | 2 | 0.25 | ns | |
| Stygobromus tenuis potomacus | 7/20/2016 | 14 | 15 | 1 | 0.62 | ns |
| 1/8/2017 | 15 | 15 | 0 | 0.00 | ns | |
| 1/8/2017 | 15 | 15 | 0 | 0.00 | ns | |
| Total | 44 | 45 | 1 | 0.61 | ns |

Number of observations of six different behaviors of Crangonyx shoemakeri, Gammarus minus, and Stygobromus tenuis potomacus in experimental and control containers. If at least one individual (and up to five) shows the behavior, this is counted as an observation. See methods for the observational protocols.
| Category | Gammarus | Crangonyx | Stygobromus | |||
|---|---|---|---|---|---|---|
| Exp | Control | Exp | Control | Exp | Control | |
| 1- walk | 3 | 20 | 19 | 138 | 40 | 55 |
| 2-swim | 3 | 125 | 1 | 54 | 5 | 53 |
| 3-burrow | 1 | 2 | 21 | 0 | 22 | 1 |
| 4-resting | 161 | 24 | 156 | 10 | 72 | 4 |
| 5-interact | 8 | 88 | 2 | 106 | 32 | 73 |
| 6-amplexus | 11 | 34 | 3 | 0 | 0 | 0 |
| Total | 189 | 293 | 202 | 308 | 171 | 186 |
When in the experimental containers, each species showed statistically significant differences in behavior relative to the control containers (Table
Log-likelihood tests for differences between behaviors in experimental and control containers for Crangonyx shoemakeri, Gammarus minus, and Stygobromus tenuis potomacus, and among control behavior and among behavior in experimental containers. n1, n2, and n3 are the total number of observations of a behavior, e.g., walking. The total number of separate observations was 180 for G. minus and C. shoemakeri; 108 for S. tenuis potomacus. For some observation periods more than one behavior was observed. See Figure
| Comparison | n1 | n2 | n3 | df | log - likelihood | p |
|---|---|---|---|---|---|---|
| G. minus-experiment vs. control2 | 188 | 291 | 4 | 147.6 | <0.01 | |
| C. shoemakeri-experiment vs. control1 | 199 | 308 | 4 | 198,9 | <0.01 | |
| S. t. potomacus-experiment vs. control1 | 171 | 186 | 4 | 70.4 | <0.01 | |
| All control | 308 | 291 | 186 | 10 | 112.1 | <0.01 |
| All experimental | 199 | 188 | 171 | 10 | 70 | <0.01 |
For C. shoemakeri, resting was also the most common activity, occurring 77% of the time (Fig.
Overall, S. tenuis potomacus showed less behavioral change than the other two species, but the differences were statistically significant compared to its control group (Table
Discussion
Adaptation to the absence of open water
Crangonyx shoemakeri and Stygobromus tenuis potomacus have evolved mechanisms allowing for their success in habitats that periodically lack open water. Both of these species exhibited similar adaptations by burrowing into the substrate during experimental drying periods. Those individuals that failed to burrow, remaining exposed on the surface, curled in ‘C’ shapes and often times lay in shallow depressions. The length of time that burrowing in clay can provide a refuge from desiccation is unknown, beyond the four week duration of the experiment. We do know that moisture is retained in clay for longer periods (see Fig.
Animals living in surface habitats that periodically dry, including many wetlands, have a large array of strategies to cope with desiccation (
In addition to its widespread occurrence in wetlands, Crangonyx is also a frequent inhabitant of caves and some of these species occur in epikarst drip pools, a subterranean habitat that occasionally dries out (Pipan and Culver 2013). Holsinger and Dickson (1976) observed burrowing behavior in an epikarst drip pool population of Crangonyx antennatus in Molly Waggle Cave in Virginia.
The only other observations of burrowing behavior in subterranean habitats comes from the European amphipod genus Niphargus.
Conservation and protection
Clay substrates provide important resources for amphipod species from nutrition to surviving environmental stressors. Our findings indicate the importance of a clay layer for Crangonyx shoemakeri and Stygobromus tenuis potomacus amphipods inhabiting hypotelminorheic habitats in the Washington, DC region. To protect these species and the endemic and endangered species, Stygobromus hayi, it is essential to maintain healthy soil. Although these cryptic species may not be found during drying periods, it is apparent that the animals are still present. Due to the difficulty in accessing the microenvironments these animals create in the soil, biological surveys may report false negatives. The application of eDNA sampling holds promise for detecting these populations (Niemiller 2017).
By 2090, it is expected that the proportion of the global land surface in extreme drought will increase tenfold from current levels (
In Australia,
In order to further assess the impact these anthropogenic changes to the environment will have on amphipod species, it is necessary to learn more about thair physiological sensitivity. To better understand amphipod adaptations and estimate the impact of warming on their fitness, further studies on the burrowing behavior of Crangonyx shoemakeri and Stygobromus tenuis potomacus should be investigated.
Acknowledgements
This research was supported by Chesapeake Watershed Cooperative Ecosystem Studies Unit Cooperative Agreement P16AC01389 between Department of the Interior, National Park Service, National Capital Parks – East and American University, “A Survey of the Fauna of Seepage Springs in National Capital Parks-East (NACE)”. All specimens were collected under CHOH-2016-SCI-0023 and NACE-2016-SCI-0002 permits. A reviewer offered many constructive comments.
References
- Abessa D, Sousa EC, Rachid BR, Mastroti RR (1998) Use of the burrowing amphipod Tiburonella viscana as a tool in marine sediments contamination assessment. Brazilian Archives of Biology and Technology 41(2): 225–230. https://doi.org/10.1590/S1516-89131998000200009
- Batzer DP, Boix D (2016) An introduction to freshwater wetlands and invertebrates. In: Batzer DP, Boix D (Eds) Invertebrates in Freshwater Wetlands.An international perspective on their ecology. Springer, Cham, 1–24. https://doi.org/10.1007/978-3-319-24978-0_1
- Batzer DP, Ruhí A (2013) Is there a core of organisms that structure macroinvertebrate assemblages in freshwater wetlands? Freshwater Biology 58(8): 1647–1659. https://doi.org/10.1111/fwb.12156
- Brooks RT (2000) Annual and seasonal variation and the effects of hydroperiod on benthic macroinvertebrates of seasonal forest (‘‘vernal’’) ponds in Central Massachusetts, USA. Wetlands 20: 707–715. https://doi.org/10.1672/0277-5212(2000)020[0707:AASVAT]2.0.CO;2
- Culver DC (1970) Analysis of simple cave communities: niche separation and species packing. Ecology 51(6): 949–958. https://doi.org/10.2307/1933622
- Culver DC, Holsinger JR, Christman MC, Pipan T (2010) Morphological differences among eyeless amphipods in the genus Stygobromus dwelling in different subterranean habitats. Journal of Crustacean Biology 30(1): 68–74. https://doi.org/10.1651/09-3156.1
- Culver DC, Holsinger JR, Feller DJ (2012) The fauna of seepage springs and other shallow subterranean habitats in the mid-Atlantic Piedmont and Coastal Plain, U.S.A. Northeastern Naturalist 19(Monograph 9): 1–42. https://doi.org/10.1656/045.019.m901
- Culver DC, Kane TC, Fong DW (1995) Adaptation and natural selection in caves: the evolution of Gammarus minus. Harvard University Press, Cambridge, 223 pp. https://doi.org/10.4159/harvard.9780674419070.c9
- Culver DC, Pipan T (2009) Biology of caves and other subterranean habitats. Oxford University Press, Oxford, 254 pp. https://doi.org/10.5860/CHOICE.47-1398
- Culver DC, Pipan T (2014) Shallow subterranean habitats: ecology, evolution, and conservation. Oxford University Press, Oxford, 258 pp. https://doi.org/10.1093/acprof:oso/9780199646173.001.0001
- DiSalvo B, Haynes J (2015) Colonization and persistence of Crangonyx gracilis (Bousfield 1958) in temporary pools. Freshwater Science 34(2): 547–554. https://doi.org/10.1086/680986
- Environment-Canada (1992) Biological test method: acute test for sediment toxicity using marine or estuarine amphipods. Report EPS1/RM/26, Ottawa, 83 pp.
- Gibert J, Culver DC (2009) Assessing and conserving groundwater biodiversity: an introduction. Freshwater Biology 54(4): 639–648. https://doi.org/10.1111/j.1365-2427.2009.02202.x
- Ginet R (1960) Ecologie, ethologie et biologie de Niphargus (Amphipodes Gammaridés hypogés). Annales de Spéléologie 15: 1–254.
- Gounot AM (1960) Recherches sur le limon argileux souterrain et sur son role nutritive pour les Niphargus. Annales de Spéléologie 15: 501–526.
- Heath RC (1983) Basic ground-water hydrology. United States Geological Survey Water Supply Paper 2220, Alexandria, 85 pp. https://doi.org/10.3133/wsp2220
- Holsinger JR (1972) The Freshwater amphipod crustaceans (Gammaridae) of North America. Biota of Freshwater Ecosystems Identification Manual No. 5, Environmental Protection Agency, Washington, DC.
- Holsinger JR, Culver DC (1970) Morphological variation in Gammarus minus Say (Amphipoda, Gammaridae), with emphasis on subterranean forms. Postilla No. 146, 24 pp.
- Holsinger JR, Dickson GW (1977) Burrowing as a means of survival in the troglobitic amphipod crustacean Crangonyx antennatus Packard (Crangonyctidae). Hydrobiologia 54(3): 195–199. https://doi.org/10.1007/BF00014284
- Jackson CR, Thompson JA, Kolka RK (2014) Wetland soils, hydrology, and geomorphology. In: Batzer DP, Sharitz RR (Eds) Ecology of freshwater and estuarine wetlands, second edition.University of California Press, Berkeley, 23–60. https://doi.org/10.1525/california/9780520247772.001.0001
- Klein E, Berg EE, Dial R (2005) Wetland drying and succession across the Kenai Peninsula Lowlands, southcentral Alaska. Canadian Journal of Forest Research 35(8): 1931–1941. https://doi.org/10.1139/x05-129
- Kundzewicz ZW, Mata LJ, Arnell NW, Doll P, Jimenez B, Miller K, Oki T, Sen Z, Shiklomanov I (2008) The implications of projected climate change for freshwater resources and their management. Hydrological Science 53(1): 3–11. https://doi.org/10.1623/hysj.53.1.3
- Marzolf RG (1965) Substrate relations of the burrowing amphipod Pontoporeia affinis in Lake Michigan. Ecology 46(5): 580–592. https://doi.org/10.2307/1934998
- Mathieu J, Turquin MJ (1992) Biological processes at the population level. II. Aquatic populations: Niphargus (stygobiont amphipod) case. In: Camacho AI (Ed.) The Natural History of Biospeleology.Monografias Museo Nacional de Ciencias Naturales, Madrid, 263–294.
- Meštrov M (1962) Un nouveau milieu aquatique souterrain: le biotope hypotelminorheique. Compte Rendus de l’Académie des Sciences, Paris 254: 2677–2679.
- Pipan T, Culver DC (2012) Wetlands in cave and karst regions. In: White WB, Culver DC (Eds) Encyclopedia of Caves.Second Edition. Elsevier/Academic Press, Amsterdam, 897–904. https://doi.org/10.1016/B978-0-12-383832-2.00129-8
- Pipan T, Fišer C, Novak T, Culver DC (2012) Fifty years of the hypotelminorheic: what have we learned? Acta Carsologica 42(2–3): 275–285. https://doi.org/10.3986/ac.v41i2-3.564
- Ponznanska M, Kakareko T, Krzyzynski M, Kobak J (2013) Effect of substratum drying on the survival and migrations of Ponto-Caspian and native gammarids (Crustacea: Amphipoda). Hydrobiologia 700(1): 47–59. https://doi.org/10.1007/s10750-012-1218-6
- Poulson TL (1964) Animals in aquatic environments: animals in caves. In: Dill DP (Ed.) Handbook of Physiology. American Physiological Society, Washington, DC.
- Reynolds JD (2016) Invertebrates of Irish turoughs. In: Batzer DP, Boix D (Eds) Invertebrates in Freshwater Wetlands.An International perspective on their ecology. Springer, Cham, 191–218. https://doi.org/10.1007/978-3-319-24978-0_6
- Robson B, Chester E, Austin C (2011) Why life history information matters: drought refuges and macroinvertebrate persistence in non-perennial streams subject to a drier climate. Marine and Freshwater Research 62(7): 801–810. https://doi.org/10.1071/MF10062
- Scheffer M (2004) Ecology of shallow lakes. Kluwer, Dordrecht, 357 pp. https://doi.org/10.1007/978-1-4020-3154-0
- Sheehyx Skeffington M, Gormally M (2007) Turloughs: a mosaic of biodiversity and management systems unique to Ireland. Acta Carsologica 36(2): 217–222. https://doi.org/10.3986/ac.v36i2.190
- Sheehy Skeffington M, Scott NE (2008) Do turloughs occur in Slovenia? Acta Carsologica 37(2–3): 291–306. https://doi.org/10.3986/ac.v37i2-3.153
- Sim LL, Davis JA, Strehlow K, McGuire M, Trayler KM, Wild S, Papas PJ, O’Connor J (2013) The influence of changing hydroregime on the invertebrate communities of temporary seasonal wetlands. Freshwater Science 32(1): 327–342. https://doi.org/10.1899/12-024.1
- Strachan SR, Chester ET, Robson BJ (2014) Microrefuges from drying for invertebrates in a seasonal wetland. Freshwater Biology 59(12): 2528–2538. https://doi.org/10.1111/fwb.12451
- Swartz EC, Deden WA, Sercu KA, Lamberson JO (1982) Sediment toxicity and the distribution of the amphipods in Commencement Bay, Washington, USA. Marine Pollution Bulletin 13(10): 259–264. https://doi.org/10.1016/0025-326X(82)90041-8
- Vanschoenwinkel B, Hulsmans A, De Roeck E, De Vries C, Seaman M, Brendonck L (2009) Community structure in temporary freshwater pools: disentangling the effects of habitat size and hydroregime. Freshwater Biology 54(7): 1487–1500. https://doi.org/10.1111/j.1365-2427.2009.02198.x
- Waterkeyn A, Grillas P, Vanschoenwinkel B, Brendonck L (2008) Invertebrate community patterns in Mediterranean temporary wetlands along hydroperiod and salinity gradients. Freshwater Biology 53(9): 1808–1822. https://doi.org/10.1111/j.1365-2427.2008.02005.x
- Wiggins GB, Mckay RJ, Smith IM (1980) Evolutionary and ecological strategies of animals in annual temporary pools. Archiv für Hydrobiologie Supplement 58: 97–206.
- Williams DD (2016) Invertebrates in groundwater springs and seeps. In: Batzer DP, Boix D (Eds) Invertebrates in freshwater wetlands.An international perspective on their ecology. Springer, Cham, 357–410. https://doi.org/10.1007/978-3-319-24978-0_11
- Williams PW (2008) The role of epikarst in karst and cave hydrology: a review. International Journal of Speleology 37(1): 1–10. https://doi.org/10.5038/1827-806X.37.1.1
- Zhang J, Holsinger JR (2003) Systematics of the freshwater amphipod genus Crangonyx in North America. Memoirs of the Virginia Museum of Natural History 6: 1–274.
- Zimmer KD, Hanson MA, Wrubleski DA (2016) Invertebrates in permanent wetlands (long-hydroperiod marshes and shallow lakes). In: Batzer DP, Boix D (Eds) Invertebrates in freshwater wetlands.An international perspective on their ecology. Springer, Cham, 251–286. https://doi.org/10.1007/978-3-319-24978-0_8